
The clustered regularly interspaced short palindromic repeats (CRISPR)-associated endonuclease 9 (CRISPR/Cas9) system has emerged as a promising technology for specific genome editing in many species. Here we constructed one vector targeting eight agronomic genes in rice using the CRISPR/Cas9 multiplex genome editing system. By subsequent genetic transformation and DNA sequencing, we found that the eight target genes have high mutation efficiencies in the T0 generation. Both heterozygous and homozygous mutations of all editing genes were obtained in T0 plants. In addition, homozygous sextuple, septuple, and octuple mutants were identified. As the abundant genotypes in T0 transgenic plants, various phenotypes related to the editing genes were observed. The findings demonstrate the potential of the CRISPR/Cas9 system for rapid introduction of genetic diversity during crop breeding.
the Agricultural Science and Technology Innovation Program of Chinese Academy of Agricultural Sciences
National Natural Science Foundation of China(31271681)
Jiangsu Agriculture Science and Technology Innovation Fund(CX(135075)
This work was supported by the National Natural Science Foundation of China (31271681, 3140101312), the Agricultural Science and Technology Innovation Program of Chinese Academy of Agricultural Sciences, and Jiangsu Agriculture Science and Technology Innovation Fund (CX(13)5075).
This article is distributed under the terms of the Creative Commons Attribution License, which permits any use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
The author(s) declare that they have no conflict of interest.
The supporting information is available online at life.scichina.com and
[1] Ashikari M., Sakakibara H., Lin S., Yamamoto T., Takashi T., Nishimura A., Angeles E.R., Qian Q., Kitano H., Matsuoka M.. Cytokinin oxidase regulates rice grain production. Science, 2005, 309: 741-745 CrossRef PubMed ADS Google Scholar
[2] Casini A., Storch M., Baldwin G.S., Ellis T.. Bricks and blueprints: methods and standards for DNA assembly. Nat Rev Mol Cell Biol, 2015, 16: 568-576 CrossRef PubMed Google Scholar
[3]
Chen
S.,
Yang
Y.,
Shi
W.,
Ji
Q.,
He
F.,
Zhang
Z.,
Cheng
Z.,
Liu
X.,
Xu
M..
[4]
Cho
Y.G.,
Eun
M.Y.,
McCouch
S.R.,
Chae
Y.A..
The semidwarf gene,
[5] Doench J.G., Fusi N., Sullender M., Hegde M., Vaimberg E.W., Donovan K.F., Smith I., Tothova Z., Wilen C., Orchard R., Virgin H.W., Listgarten J., Root D.E.. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat Biotechnol, 2016, 34: 184-191 CrossRef PubMed Google Scholar
[6] Fan C., Xing Y., Mao H., Lu T., Han B., Xu C., Li X., Zhang Q.. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet, 2006, 112: 1164-1171 CrossRef PubMed Google Scholar
[7] Fukuma M., Ganmyo Y., Miura O., Ohyama T., Shimizu N.. Cloning and characterization of a human genomic sequence that alleviates repeat-induced gene silencing. PLoS One, 2016, 11: e0153338 CrossRef PubMed ADS Google Scholar
[8]
Generoso
W.C.,
Gottardi
M.,
Oreb
M.,
Boles
E..
Simplified CRISPR-Cas genome editing for
[9]
Hiei
Y.,
Ohta
S.,
Komari
T.,
Kumashiro
T..
Efficient transformation of rice (
[10] Hu X., Wang C., Fu Y., Liu Q., Jiao X., Wang K.. Expanding the range of CRISPR/Cas9 genome editing in rice. Mol Plant, 2016, 9: 943-945 CrossRef PubMed Google Scholar
[11]
Huang
X.,
Qian
Q.,
Liu
Z.,
Sun
H.,
He
S.,
Luo
D.,
Xia
G.,
Chu
C.,
Li
J.,
Fu
X..
Natural variation at the
[12] Jiang L., Liu X., Xiong G., Liu H., Chen F., Wang L., Meng X., Liu G., Yu H., Yuan Y., Yi W., Zhao L., Ma H., He Y., Wu Z., Melcher K., Qian Q., Xu H.E., Wang Y., Li J.. DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature, 2013, 504: 401-405 CrossRef PubMed ADS Google Scholar
[13]
Li
M.,
Li
X.,
Zhou
Z.,
Wu
P.,
Fang
M.,
Pan
X.,
Lin
Q.,
Luo
W.,
Wu
G.,
Li
H..
Reassessment of the four yield-related genes
[14] Li T., Huang S., Zhao X., Wright D.A., Carpenter S., Spalding M.H., Weeks D.P., Yang B.. Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes. Nucleic Acids Res, 2011, 39: 6315-6325 CrossRef PubMed Google Scholar
[15] Liu X., Zhang Y., Cheng C., Cheng A.W., Zhang X., Li N., Xia C., Wei X., Liu X., Wang H.. CRISPR-Cas9-mediated multiplex gene editing in CAR-T cells. Cell Res, 2017, 27: 154-157 CrossRef PubMed Google Scholar
[16] Ma X., Liu Y.G.. CRISPR/Cas9-based multiplex genome editing in monocot and dicot plants. Curr Protoc Mol Biol, 2016, 115: 31.6.1-31.6.21 CrossRef Google Scholar
[17] Ma X., Zhu Q., Chen Y., Liu Y.G.. CRISPR/Cas9 platforms for genome editing in plants: developments and applications. Mol Plant, 2016, 9: 961-974 CrossRef PubMed Google Scholar
[18] Ma X., Chen L., Zhu Q., Chen Y., Liu Y.G.. Rapid decoding of sequence-specific nuclease-induced heterozygous and biallelic mutations by direct sequencing of PCR products. Mol Plant, 2015a, 8: 1285-1287 CrossRef PubMed Google Scholar
[19] Ma X., Zhang Q., Zhu Q., Liu W., Chen Y., Qiu R., Wang B., Yang Z., Li H., Lin Y., Xie Y., Shen R., Chen S., Wang Z., Chen Y., Guo J., Chen L., Zhao X., Dong Z., Liu Y.G.. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol Plant, 2015b, 8: 1274-1284 CrossRef PubMed Google Scholar
[20] Mao H., Sun S., Yao J., Wang C., Yu S., Xu C., Li X., Zhang Q.. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc Natl Acad Sci USA, 2010, 107: 19579-19584 CrossRef PubMed ADS Google Scholar
[21] Mitsuda S.H., Shimizu N.. Epigenetic repeat-induced gene silencing in the chromosomal and extrachromosomal contexts in human cells. PLoS One, 2016, 11: e0161288 CrossRef PubMed ADS Google Scholar
[22] Miura K., Ikeda M., Matsubara A., Song X.J., Ito M., Asano K., Matsuoka M., Kitano H., Ashikari M.. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nat Genet, 2010, 42: 545-549 CrossRef PubMed Google Scholar
[23]
Monna
L.,
Kitazawa
N.,
Yoshino
R.,
Suzuki
J.,
Masuda
H.,
Maehara
Y.,
Tanji
M.,
Sato
M.,
Nasu
S.,
Minobe
Y..
Positional cloning of rice semidwarfing gene,
[24] Patron N.J.. DNA assembly for plant biology: techniques and tools. Curr Opin Plant Biol, 2014, 19: 14-19 CrossRef PubMed Google Scholar
[25]
Piao
R.,
Jiang
W.,
Ham
T.H.,
Choi
M.S.,
Qiao
Y.,
Chu
S.H.,
Park
J.H.,
Woo
M.O.,
Jin
Z.,
An
G.,
Lee
J.,
Koh
H.J..
Map-based cloning of the
[26] Qi W., Zhu T., Tian Z., Li C., Zhang W., Song R.. High-efficiency CRISPR/Cas9 multiplex gene editing using the glycine tRNA-processing system-based strategy in maize. BMC Biotechnol, 2016, 16: 58 CrossRef PubMed Google Scholar
[27] Sakuma T., Masaki K., Abe-Chayama H., Mochida K., Yamamoto T., Chayama K.. Highly multiplexed CRISPR-Cas9-nuclease and Cas9-nickase vectors for inactivation of hepatitis B virus. Genes Cells, 2016, 21: 1253-1262 CrossRef PubMed Google Scholar
[28] Shan Q., Wang Y., Li J., Gao C.. Genome editing in rice and wheat using the CRISPR/Cas system. Nat Protoc, 2014, 9: 2395-2410 CrossRef PubMed Google Scholar
[29] Shan Q., Wang Y., Li J., Zhang Y., Chen K., Liang Z., Zhang K., Liu J., Xi J.J., Qiu J.L., Gao C.. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat Biotechnol, 2013, 31: 686-688 CrossRef PubMed Google Scholar
[30] Shen L., Wang C., Fu Y., Wang J., Liu Q., Zhang X., Yan C., Qian Q., Wang K.. QTL editing confers opposing yield performance in different rice varieties. J Integr Plant Biol, 2016, in press: doi: 10.1111/jipb.12501 CrossRef PubMed Google Scholar
[31] Si L., Chen J., Huang X., Gong H., Luo J., Hou Q., Zhou T., Lu T., Zhu J., Shangguan Y., Chen E., Gong C., Zhao Q., Jing Y., Zhao Y., Li Y., Cui L., Fan D., Lu Y., Weng Q., Wang Y., Zhan Q., Liu K., Wei X., An K., An G., Han B.. OsSPL13 controls grain size in cultivated rice. Nat Genet, 2016, 48: 447-456 CrossRef PubMed Google Scholar
[32] Song X.J., Huang W., Shi M., Zhu M.Z., Lin H.X.. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet, 2007, 39: 623-630 CrossRef PubMed Google Scholar
[33] Sood, B.C., and Sidiq, E.A. (1978). A rapid technique for scent determination in rice. Indian J Genet Plant Breed 38, 268?275. Google Scholar
[34]
Spielmeyer
W.,
Ellis
M.H.,
Chandler
P.M..
Semidwarf (
[35] Wang C., Shen L., Fu Y., Yan C., Wang K.. A Simple CRISPR/Cas9 system for multiplex genome editing in rice. J Genets Genomics, 2015a, 42: 703-706 CrossRef PubMed Google Scholar
[36] Wang S., Li S., Liu Q., Wu K., Zhang J., Wang S., Wang Y., Chen X., Zhang Y., Gao C., Wang F., Huang H., Fu X.. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat Genet, 2015b, 47: 949-954 CrossRef PubMed Google Scholar
[37] Wang X., Niu Y., Zhou J., Yu H., Kou Q., Lei A., Zhao X., Yan H., Cai B., Shen Q., Zhou S., Zhu H., Zhou G., Niu W., Hua J., Jiang Y., Huang X., Ma B., Chen Y.. Multiplex gene editing via CRISPR/Cas9 exhibits desirable muscle hypertrophy without detectable off-target effects in sheep. Sci Rep, 2016, 6: 32271 CrossRef PubMed ADS Google Scholar
[38] Wood A.J., Lo T.W., Zeitler B., Pickle C.S., Ralston E.J., Lee A.H., Amora R., Miller J.C., Leung E., Meng X., Zhang L., Rebar E.J., Gregory P.D., Urnov F.D., Meyer B.J.. Targeted genome editing across species using ZFNs and TALENs. Science, 2011, 333: 307-307 CrossRef PubMed ADS Google Scholar
[39] Wu X., Tang D., Li M., Wang K., Cheng Z.. Loose plant architecture1, an INDETERMINATE DOMAIN protein involved in shoot gravitropism, regulates plant architecture in rice. Plant Physiol, 2013, 161: 317-329 CrossRef PubMed Google Scholar
[40] Xing H.L., Dong L., Wang Z.P., Zhang H.Y., Han C.Y., Liu B., Wang X.C., Chen Q.J.. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol, 2014, 14: 327 CrossRef PubMed Google Scholar
[41]
Yano
M.,
Katayose
Y.,
Ashikari
M.,
Yamanouchi
U.,
Monna
L.,
Fuse
T.,
Baba
T.,
Yamamoto
K.,
Umehara
Y.,
Nagamura
Y.,
Sasaki
T..
[42]
Yu
H.,
Murchie
E.H.,
González-Carranza
Z.H.,
Pyke
K.A.,
Roberts
J.A..
Decreased photosynthesis in the
[43]
Zhang
Z.,
Mao
Y.,
Ha
S.,
Liu
W.,
Botella
J.R.,
Zhu
J.K..
A multiplex CRISPR/Cas9 platform for fast and efficient editing of multiple genes in
[44]
Zhou
Y.,
Zhu
J.,
Li
Z.,
Yi
C.,
Liu
J.,
Zhang
H.,
Tang
S.,
Gu
M.,
Liang
G..
Deletion in a quantitative trait gene
Figure 1
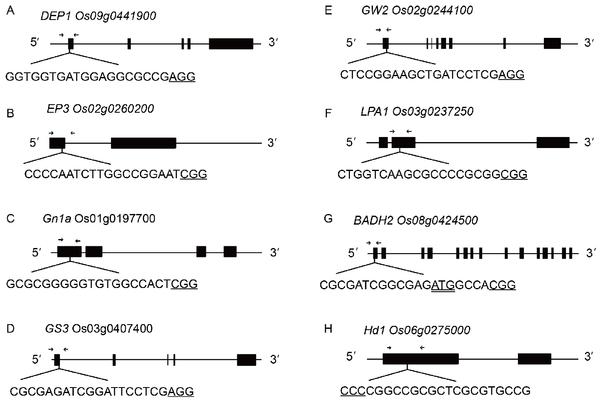
Schematic diagram of the targeted sites in eight genes. A–H, The targeted sites are labeled in black uppercase letters. The initiation codons are underlined twice. The protospacer adjacent motif (PAM) sequences are underlined once. The arrows show the regions around the editing sites.
Figure 2
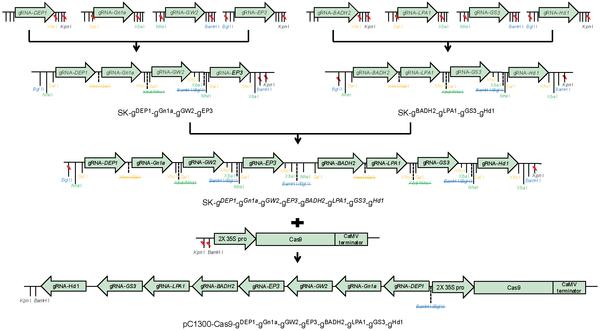
Flow diagram of an octuple CRISPR/Cas9 system for multiplex gene editing in rice. The restriction sites used for cloning are labeled.
Figure 3
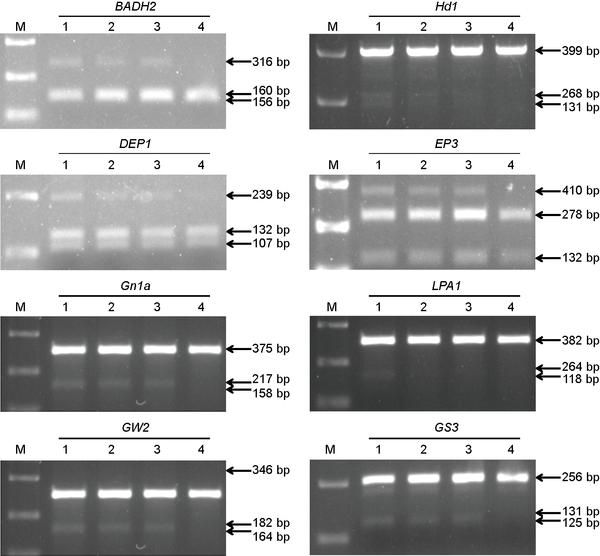
PCR/RE assay of mutations at eight loci in rice protoplast.
Figure 4
Characterization of targeted editing in T0 rice plants. A, Editing efficiencies of eight agronomic genes in T0 plants. B, Editing efficiencies of wild type (WT), homozygous mutations, and heterozygous mutations in each gene. C, Numbers of plants with different mutation genes. D, Editing efficiencies of off-target genes in T0 plants.
Figure 5
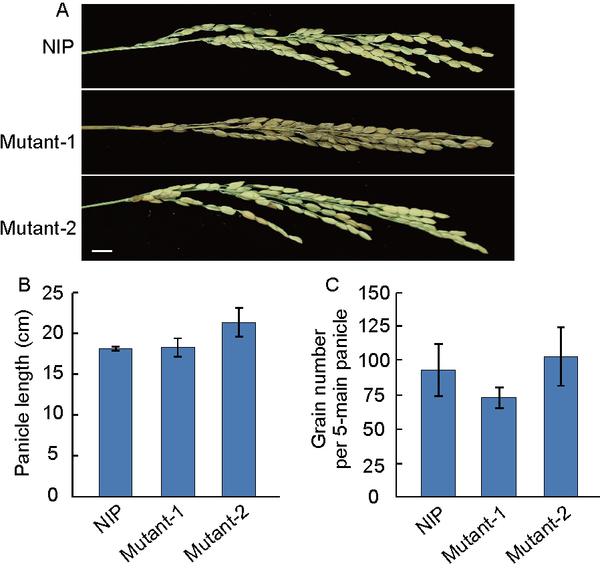
Comparison of panicle traits among Nipponbare (NIP), Mutant 1, and Mutant 2. A, The morphology of the panicles of the NIP, Mutant 1, and Mutant 2. Scale bar, 1 cm. B, Comparison of panicle length among NIP, Mutant 1, and Mutant 2. C, Comparison of grain number per panicle among NIP, Mutant 1, and Mutant 2. Values in B and C are means±standard deviations (SD),
Figure 6
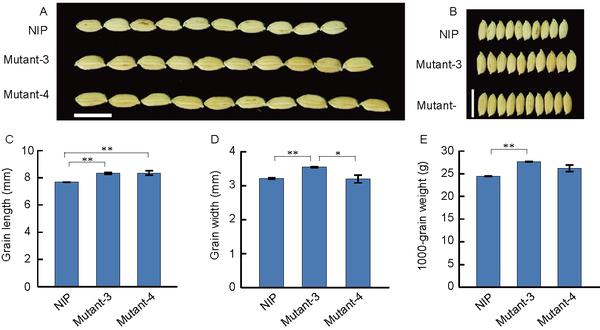
Comparison of seed size among Nipponbare (NIP), Mutant 3, and Mutant 4. Aand B, Grain shape of the NIP, Mutant 3, and Mutant 4. Scale bar, 1 cm. C, Comparison of grain length among NIP, Mutant 3, and Mutant 4. D, Domparison of grain width among NIP, Mutant 3, and Mutant 4. E, Comparison of 1,000-grain weight among NIP, Mutant 3, and Mutant 4. Values in C, D, and F are means±standard deviation (s.d.),
Figure 7
Phenotypes of plants with different gene combinations. A–F, Nipponbare (NIP). The letters (a–h), up from the plants represent
Type of gene mutation |
Genotype |
No. of plants |
Sum |
Double mutations |
AABBccDdEEFFGGHH |
3 |
3 |
Quintuple mutations |
AabbccddEEffGGHH |
2 |
8 |
aaBbCcddEEffGGHH |
1 |
||
aaBBccddEEFFGgHh |
2 |
||
AABbccddEEffGgHH |
1 |
||
AABbccddEEffGGHh |
1 |
||
AABBCcDdeeffGGHh |
1 |
||
Sextuple mutations |
aabbCcddeeffGGHH |
1 |
6 |
aaBbCcddeeffGGHH |
2 |
||
aabbccddeeFfGGHH |
1 |
||
aabbccddeeffGGHH |
1 |
||
AaBbCCddEeFfGgHH |
1 |
||
Septuple mutations |
aabbccddEEffGgHh |
1 |
10 |
aaBbccddEEFfGgHh |
1 |
||
aabbccddeeFFGghh |
2 |
||
aaBbccddEeFFGgHh |
1 |
||
aabbccddeeFfGGhh |
1 |
||
aabbccddeeffGGhh |
1 |
||
aaBbccddeeFfGGhh |
2 |
||
aabbccddeeffGGHh |
1 |
||
Octuple mutations |
AaBbccddEeFfGgHh |
1 |
9 |
aabbccddeeFfGghh |
4 |
||
aaBbccddEeFfGgHh |
1 |
||
Aabbccddeeffgghh |
2 |
||
AabbccddeeFfGgHh |
1 |
a)The
letters (a–h), represent

Copyright 2019 Science China Press Co., Ltd. 科学大众杂志社有限责任公司 版权所有
京ICP备18024590号-1

 Download PDF
Download PDF


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}